- Study of internal structure of the body of an organism is known as Anatomy.
- Higher plants have highly complex body made up of different types of cells. All cells have the same origin but afterwards they get differentiated into different types of cells.
- A group of similar or dissimilar cells having common origin and performing a specific function is known as tissue.
Tissues are mainly divided into two categories:
(1) Meristematic tissues or Meristems
(2) Permanent tissue
1. MERISTEMATIC TISSUES OR MERISTEMS
- The word “Meristem” originated from “Meristos” (Greek = divided) was coined by Nageli (1858). A group of cells which are more active and capable of showing continuous division and redivisions, is called as meristematic tissue.
- In plants growth is restricted to a particular region, where meristem is present.
Characteristics of Meristems
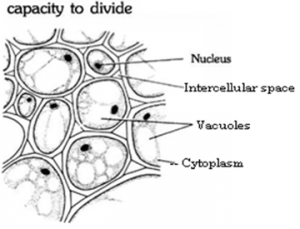
- Cells of meristems are living and are at active divisional stage.
- The cells are usually small and isodiametric. The cells of cambium are, however, elongated.
- The cells are compactly arranged without any intercellular spaces.
- The cell wall is thin and made up of cellulose.
- Cytoplasm is dense, granular with a central, large and prominent nucleus.
- Vacuoles are either very small or absent. The cells of cambium are, however, vacuolated.
- Metabolic activity is high, so is the rate of respiration.
- Cytoplasm has all the cell organelles, but instead of plastids there are proplastids (eoplasts). Ribosomes are abundant.
Types of meristems
The meristems may be classified on the basis of their mode of origin, position etc.:
(i) According to origin and development : On the basis of origin, meristematic tissues are of three types:
(a) Promeristem or Primordial meristem : The promeristem originates from embryo and therefore, called primordial or embryonic meristem. It is present in the regions where an organ or a part of plant body is initiated. A group of initial cells that lay down the foundation of an organ or a plant part, is called promeristem. This group consists of limited number of cells, which divide repeatedly to give rise to primary meristem. The promeristem gives rise to all other meristems including the primary meristem.
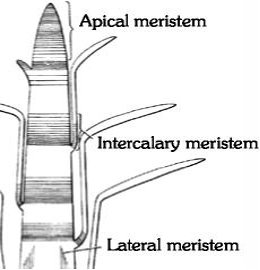
(b) Primary Meristem: A primary meristem originates from promeristem and retains its meristematic activity. It is located in the apices of roots, stems and the leaf primordial, e.g., Apical meristem and Intercalary meristem.
(c) Secondary Meristem: The secondary meristems are so called because they originate from permanent cells. E.g., phellogen or cork cambium, is an important example of secondary meristem. The secondary meristems produce secondary tissues in the plant body and add new cells for effective protection and repair. They occur in mature regions of roots and shoots.
(ii) According to position : On the basis of their position in the plant body meristems are classified into three categories :
Various meristamatic tissue
(a) Apical meristem : This meristem is located at the growing apices of main, as well as lateral shoots and roots. These cells are responsible for linear growth of an organ. Position of apical cells may either be strictly terminal or subterminal.
(b) Intercalary meristem : These are the portions of apical meristems which are separated from the apex during the growth of axis and formation of permanent tissues. (e.g., Mint), base of internode (e.g., stem of many monocots viz., Wheat, Grasses, Pteridophyets like Equisetum). It occur between mature tissues.
(c) Lateral meristem : These meristems occur laterally in the axis, parallel to the sides of stems and roots. The cambium of vascular bundles (Fascicular, interfascicular and extrastelar cambium) and the cork cambium or phellogen belong to this category and are found in dicotyledons and gymnosperms resulting in an increase in their diameter.
Differences between apical and lateral meristems
| S.No. | Apical meristem | Lateral meristem |
| 1. | It occurs at the apex of stem, root and their branches. | It is found in lateral position parallel to circumference of the organs. |
| 2. | It is a primary meristem. | It is secondary meristem, except intra-fascicular cambium. |
| 3. | Cells divide in different planes. | Cells divide in one plane patricianly both on the outer and inner sides. |
| 4. | It produces primary tissues. | It gives rise to secondary tissues. |
| 5. | It brings about growth in length. | It causes growth in girth and thickness. |
Structure and Organisation of Apical Meristem
(i) Vegetative shoot apex: Shoot apex was first recognized by Wolff (1759). Shoot apex is derived from meristem present in the plumule of embryo and occurs at the tip of stem and its branches as terminal bud. It also occurs in the inactive state in the axils of leaves as lateral buds. The tip of the shoot apex is dome-shaped and from it flanks, at the base of the dome, one or more leaf primordia. This continues throughout the vegetative phase. Many theories have been put forward to explain shoot apex, such as :
(a) Apical Cell theory: This theory was proposed by Nageli (1858). According to this theory, shoot apical meristem consists of single apical cell. This theory is applicable in case of higher algae, bryophytes and in many pteridophytes but not in higher plants (i.e., gymnosperms and angiosperms).
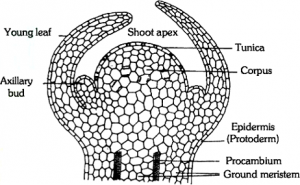
vegetative shoot apex
(b) Histogen theory: It was proposed by Hanstein (1870). According to this theory, the shoot apical meristem consists of three distinct meristematic zones or layers (or histogens).
- Dermatogen: Outermost layer and it forms epidermis and epidermal tissue system.
- Periblem: It is the middle layer that gives rise to cortex and endodermis.
- Plerome: Innermost layer forms pith and stele.
(c) Tunica corpus theory: This theory was proposed by Schmidt (1924). According to this theory, the shoot apex consists of two distinct zones.
- Tunica: It is mostly single layered and forms epidermis. The cells of tunica are smaller than corpus. The tunica shows only anticlinal division and is responsible for surface growth.
- Corpus: It represents the central core with larger cells. Corpus shows divisions in all planes and it is responsible for volume growth.
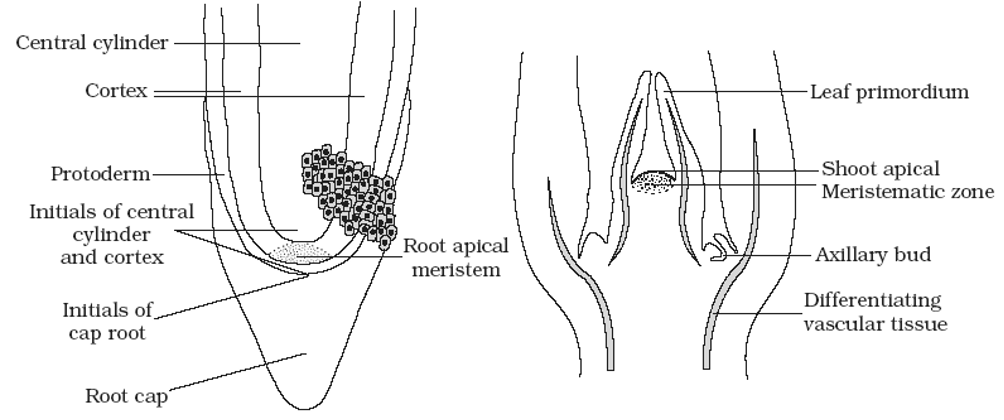
(ii) Root apex: A group of initial cells, present at the subterminal region of the growing root tip, which is protected by a root cap is called root apical meristem or root apex. It is embryonic in origin and formed from the radicle part of embryo. However, in adventitious roots it is produced from derivatives of root apex. The root apex differs from shoot apex as it is short and more or less uniform due to complete absence of lateral appendages (leaves and branches) and differentiation of nodes and internodes. According to Hanstein (1870) root apex of most of the dicotyledons also consists of three meristematic zones – plerome, periblem and dermatogen (fourth meristem calyptrogen to form root cap). Regarding the apical organisation of root following theories have been put forward.
(a) Korper -Kappe theory: It was proposed by Schuepp (1917). This theory is comparable with the tunica and corpus theory of shoot apex. Korper means body and Kappe means cap.
(b) Quiescent centre theory: It was proposed by Clowes (1961). According to him, in addition to actively dividing cells, a zone of inactive cells are present in the central part of the root apex, called quiescent centre. The cells in this region have light cytoplasm, small nuclei, lower concentration of DNA, RNA and protein. These cells also contain fewer number of mitochondria, less endoplasmic reticulum and small dictyosomes.
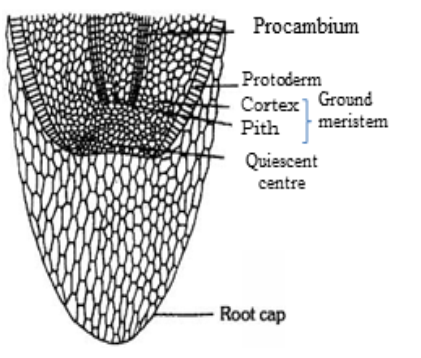
Root apical meristem
Permanent Tissues
Permanent tissues are made up of mature cells which develops due to division and differentiation in meristematic tissues. The cells of these tissues are either living or dead, thin-walled or thick-walled. Permanent tissues are of three types :
PERMANENT TISSUES
2. SIMPLE TISSUES
Simple tissues are a group of cells which are all alike in origin, form and function. They are further grouped under three categories :
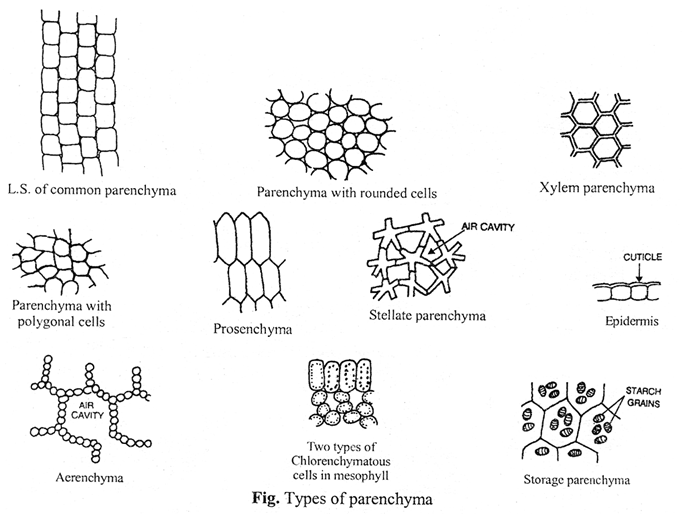
Parenchyma
Parenchyma is most simple and unspecialized tissue which is concerned mainly with the vegetative activities of the plant. The main characteristics of parenchyma cells are :
(a) The cells are thin-walled and soft.
Parenchyma in T.S.
(b) The cells usually are living and possess a distinct nucleus.
(c) The cells contain well-developed intercellular spaces amongst them.
(d) The cytoplasm is vacuolated and cell wall is made up of cellulose.
(e) The shape may be oval, spherical, cylindrical, rectangular or stellate (star shaped) as in leaf petioles of banana, canna and some hydrophytes.
(f) This tissue is generally present in almost all the organs of plants, i.e., roots, stems, leaves, flowers, fruits and seeds.
(g) If they enclose large air spaces they are called as aerenchyma; if they develop chlorophyll, they are called as chlorenchyma and if they are elongated cells with tapering ends, they are called as prosenchyma.
Functions: They perform the following functions :
- Storage of food materials. g., Carrot, Beetroot etc.
- Chlorenchyma helps in photosynthesis.
- Aerenchyma helps in floating of the aquatic plants (Hydrophytes) and also help in gaseous exchange during respiration and photosynthesis, g., Hydrilla.
- In turgid state they give rigidity to the plant organ.
- In emergency they behave like meristematic cells and help in healing of the various plant injuries.
- Sometimes they store secretory substances (ergastic substance) such as tannins, resins and gums and they called as idioblasts.
Collenchyma
It is the tissue of primary body. Occurs in layer below epidermis in dicotyledon stem. Present as a continuous layer or in patches.
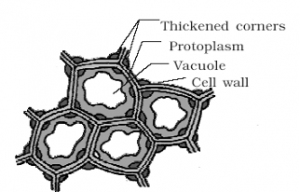
Collenchyma
- The cells of this tissue contain protoplasm and are living.
- The cell walls are thickened at the corners and are made up of cellulose, hemicellulose and pectin.
- They are compactly arranged cells, oval, spherical or polygonal in outline.
- No intercellular spaces are present.
- The tissue is extensible and have capacity to expand.
- These cells assimilate food when they contain chloroplast.
- They provide mechanical strength to younger part where xylem is less developed, such as young stem, petiole of leaf etc.
- They are usually absent in monocots stems and in roots.
Differences between parenchyma and collenchyma
| S. No. | Parenchyma | Collenchyma |
| 1. | This is found everywhere in the plant body. | It is found in hypodermis of dicot stem and at the leaf base. |
| 2. | It can be both primary and secondary tissue. | It is usually a primary tissue. |
| 3. | Cells are isodiametric. | Cells are elongated. |
| 4. | It provides turgidity to softer tissues. | It provides mechanical strength as well as elasticity to soft structures. |
| 5. | This can dedifferentiate rapidly to form meristematic cells. | Cells dedifferentiate only rarely. |
Sclerenchyma
The main features of sclerenchyma are :
- It consist of thick-walled dead cells.
- The cells vary in shape, size and origin.
- They possess hard and extremely thick secondary walls due to uniform deposition of lignin.
- In the beginning, the cells are living and have protoplasm but due to deposition of impermeable secondary walls they become dead.
Types of sclerenchyma: They are of two types :
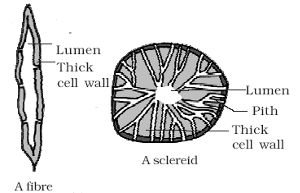
(a) Sclerenchymatous fibres :
- These are greatly elongated and tapering at both the ends.
- The fully developed fibre cells are always dead. They are polygonal in transverse section and walls are highly lignified.
- Intercellular spaces are absent and lumen is highly obliterated. The walls show simple and oblique pits.
Sclerenchyma
- They provide mechanical strength to the plant.
- Some of the longest fibre yielding plants are Linum usitatissimum (Flax or Alsi), Corchorus, Cannabis, etc.
- The fibres are present in hypodermis of monocot stem, in pericycle of many dicots, in secondary wood and vascular bundle sheath in monocot stems.
Types of Fibres
- On the basis of source and economic use, fibres are of following types:
- Surface fibres: Fibres obtained from seed coat (testa) of cotton and mesocarp of coconut (coir of commerce) are surface fibres. Cotton fibres are of two types, lint and fuzz. The lint is longer and economically useful while the fuzz is shorter and is not useful.
- Bast fibres: Fibres obtained from phloem and pericycle are bast or bass fibres. They are most exploited economically, e.g., Jute, Flax, Hemp.
- Xylary or wood fibres: Fibres associated with xylem are called xylary fibres. Two types of xylary fibres are recognized, libriform fibres and fibre tracheids. Libriform fibres are longer, thick walled and with simple pits while fibre tracheids are shorter, less thickened usually with bordered pits.
(b) Stone cells or Sclereids:
- They are lignified, extremely thick walled so that the lumen of the cells is almost obliterated. Cells may be spherical, oval, cylindrical, T-shaped or even stellate.
- They are generally found in hard parts of the plant, e.g., endocarp of Walnut and Coconut.
- They form part of seed coat in some members of leguminosae. The sclereids provide mechanical support and hardness to the soft parts. Sclereids may be :
▪ Brachysclereids or stone cells: These are small and more or less isodiametric in shape. They occur in the cortex, pith, phloem, and pulp of fruits (e.g., Pyrus).
▪ Macrosclereids or rod cells: These are rod-shaped elongated sclereids usually found in the leaves, cortex of stem and outer seed coats. E.g.,Legumes. - Osteosclereids: These are bone or barrel-shaped sclereids dilated at their ends. e.g., leaf of Hakea.
- Astrosclereids or stellate cells : These are star-shaped sclereids with extreme lobes or arms, e.g., leaf of Nymphaea.
- Trichosclereids or internal hairs : These are hair-like sclereids found in the intercellular spaces in the leaves and stem of some hydrophytes. E.g., Fig leaf
3. COMPLEX TISSUES
A group of more than one type of cells having common origin and working together as a unit, is called complex permanent tissue. The important complex tissues in vascular plants are : xylem and phloem. Both these tissues together are called as vascular tissue.
Xylem
The term xylem was introduced by Nageli (1858). Xylem is a conducting tissue which conducts water and mineral nutrients upwards from the roots to the leaves.
It also provides mechanical strength to plant parts. On the basis of origin xylem is of two types
Types of Xylem
- Primary xylem : It is derived from procambium during primary growth. It consists of protoxylem and metaxylem.
- Secondary xylem : It is formed from vascular cambium during secondary growth.
Xylem Cells
Xylem is composed of four types of cells
(a) Tracheids: The tracheids are elongated tube like cells with tapering or rounded or oval ends with hard and lignified walls.
- All the vascular plants have tracheids in their xylem.
- The walls are not much thickened.
- The cells are without protoplast and are dead on maturity. The tracheids of secondary xylem have fewer sides and are more sharply angular than the tracheids of primary xylem.
- The cell cavity or lumen of a tracheid is large and without any contents. Tracheids possess bordered pits. Maximum bordered pits are formed in gymnospermous tracheids.
- They also possess various kinds of thickenings, e.g., annular, spiral, scalariform, reticulate or pitted tracheids.
- The main function of tracheids is to conduct water and minerals from the root to the leaf. They also provide strength and mechanical support to the plant.
(b) Xylem Vessels: Vessels are rows of elongated tube-like cells, placed end to end with their end walls dissolved.
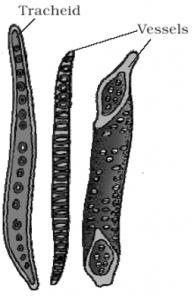
- Vessels are multicellular with wide lumen.
- The vessels may be classified into several types according to the thickening developed in their wall. They may be annular, spiral, scalariform, reticulate or pitted.
- Vessels are absent in pteridophytes and gymnosperms (except Ephedra, Gnetum, Selaginella, Pteridium).
- In angiosperms (porous wood) vessels are always present (Vessels are absent in family – Winteraceae and Trochodendraceae of Angiosperm i.e., Wintera, Trochodendron).
- Vessels along with tracheids form the main tissue of xylem vascular bundles of the angiosperms and help in conduction.
- It also provides mechanical support to the plant.
- Vessel members are interconnected through perforations in their common wall.
(c) Wood (xylem) parenchyma:
- These are the living parenchymatous cells, associated with xylem therefore known as wood parenchyma.
- They serve as storage of reserve food and also help in conduction of water upwards through tracheids and vessels.
- Their cell wall is cellulosic, radial conduction of water takes place through ray parenchyma.
- Radial conduction of water takes place through ray parenchyma.
(d) Wood (xylem) fibres:
- The long, slender, pointed, dead and sclerenchymatous cells found associated with xylem are termed wood fibres.
- They possess mostly thickened walls and few small pits. These pits are found abundantly in woody dicotyledons.
- They provide mechanical strength to xylem and various other organs of plant body.
Phloem (blast)
Term “Phloem” was given by Nageli. Its main function is the transport of organic food materials from leaves to stem and roots in a downward direction. On the basis of origin phloem is of two types:
Types of Phloem
(a) Primary phloem: It is formed by procambium during primary growth.
- It may or may not show differentiation into protophloem (consists of sieve elements and parenchyma) and metaphloem (develop after protophloem and consists of sieve elements, parenchyma and fiber).
- During the primary growth the protophloem elements are crushed by the surrounding tissues and disappear. This process is known as obliteration
- Primary phloem consists of sieve elements, parenchyma and fibre.
(b) Secondary Phloem: It is produced during secondary growth by vascular cambium.
It consists of the following elements :
- Sieve elements
- Companion cells
- Phloem parenchyma
- Phloem fibres or bast fibres
(1) Sieve element
(i) They are long tube-like cells placed end to end, forming a continuous channel in the plant parts.
(ii) Their cell wall is made up of cellulose.
(iii) Their transverse wall is perforated like a normal sieve and hence they are called as sieve tubes.
(iv) Nucleus is not found in these cells.
(v) Each sieve tube has a lining of cytoplasm near its periphery.
(vi) Their main function is to translocate the food material from one part to the other.
(2) Companion cell
(i) They are thin-walled cells which are associated with sieve tubes.
(ii) They are more or less elongated.
(iii) They are connected with the sieve tube through sieve pore.
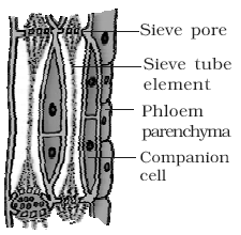
(iv) They contain nucleus and are therefore, living in nature. This nucleus controls functioning of sieve tube.
(v) They are not found in pteridophytes and gymnosperms but are always present in angiosperms.
(vi) Companion cells helps in maintaining pressure gradient in sieve tube.
(3) Phloem parenchyma : The parenchyma associated with the phloem is called phloem parenchyma. The cells are elongated with rounded ends and possess cellulosic cell walls. These cells are living and store food reserves in the form of starch and fats. They are present in pteridophytes and most of dicotyledonous angiosperms. They are absent in monocots.
(4) Phloem or Bast fibres: The sclerenchymatous fibres associated with the phloem are called as phloem fibres. These are also known as bast fibres. The fibres are elongated lignified cells with simple pits. The ends of these cells may be pointed, needle like or blunt. They are non-living cells that provide mechanical support to the organs.
Differences between Xylem and Phloem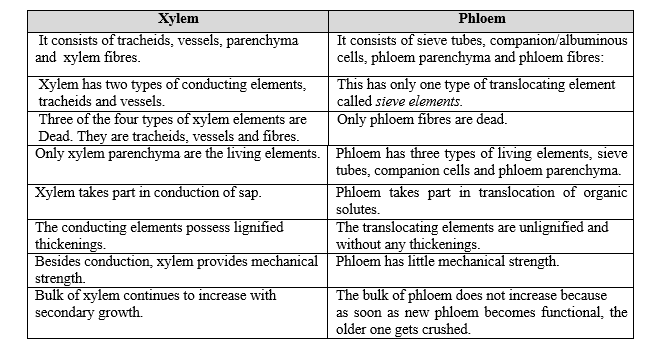
SECRETORY TISSUES
These tissue perform special function in plants, e.g., secretion of resins gum, oil and latex.
These tissues are of two types :
(1) Laticiferous tissues
(2) Glandular tissues
Laticiferous tissues
They are made up of thin walled, elongated, branched and multinucleate (coenocytic) structures that contain colourless, milky or yellow coloured juice called latex. These occur irregularly distributed in the mass of parenchymatous cells. latex is contained inside the laticiferous tissue which is of two types :
(i) Latex cell: A laticiferous cell is a very highly branched cell with long slender processes, ramifying in all directions in the ground tissue of the organ.
- They do not fuse and form a network.
- Plants having such tissues are called simple or non-articulated laticifers. e.g., Calotropis (Asclepiadaceae) Nerium, Vinca (Apocyanaceae), Euphorbia (Euphorbiaceae), Ficus (Moraceae).
(ii) Latex vessels : They are formed due to fusion of cells and form network like structure in all directions. At maturity, they form a highly ramifying system of channels full of latex inside the organ. Plants having such tissues are called compound or articulated laticifers. e.g., Argemone, Papaver (Papaveraceae), Sonchus (Compositae), Hevea, Manihot (Euphorbiaceae).
Glandular tissue
This is a highly specialized tissue consisting of glands, discharging diverse functions, including secretory and excretory. Glands may be external or internal.
(i) External glands : They generally occur on the epidermis of stem and leaves as glandular hair as in Plumbago and Boerhaavia, stinging hair secrete poisonous substance in Urtica, nectar secreting glands in flowers or leaves, e.g., Rutaceae and Euphorbiaceae. Digestive enzyme secreting glands in insectivorous plants, e.g., Drosera (Sundew), Nepenthes (Pitcher plant).
(ii) Internal glands: These are present internally and are of several types, e.g., oil glands in Citrus and Eucalyptus, resinous ducts in Pinus, mucilage canals in Cycas. Water secreting glands (hydathodes) in Colocasia (present at the tip of leaves), Tropaeoleum (along margin), etc. The glands which secrete essential oil are called osmophores (osmotrophs).
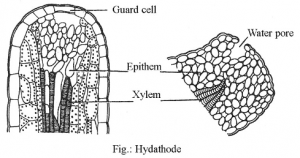
5. HYDATHODES
- They are the water exuding structures of plants inhabiting the humid tropics.
- These hydathodes are found in the leaves of many angiosperms and are located on leaf margins (e.g., Tropaeolum) or at the tip (e.g.. Colocasia).
- The cells of the mesophyll adjacent to the vascular bundle proliferate to give rise to epithem. The cells of the epithem are thin walled, elongated with dense cytoplasm and deficient in chloroplasts. They have a well-developed system of intercellular spaces and are in close contact with terminal tracheary elements.
- Overlying the epithem are present two guard cells. The water that moves out of the tracheids under conditions of high root pressure and humidity is ultimately discharged through the terminal pore of the hydathode.
6. THE TISSUE SYSTEM
The various types of tissues present in the body of a plant perform different functions. Several tissues may collectively perform the same function. A collection of tissues performing the same general function is known as a “Tissue System”. There are three major tissue systems present in plants.
(1) Epidermal tissue system
(2) Ground or fundamental tissue system
(3) Vascular or conducting tissue system
Epidermal tissue system
The tissues of this system originate from the outermost layer of apical meristem. It forms the outermost covering of various plant organs which remains in direct contact with the environment.
♦ Epidermis: Epidermis is composed of single layer of cells.
- These cells vary in their shape and size and form a continuous layer interrupted by stomata. In some cases epidermis may be multilayered, e.g., Ficus, Nerium, Peperomia, Begonia etc.
- The epidermal cells are living, parenchymatous, and compactly arranged without intercellular spaces.
- Certain epidermal cells of some plants or plant parts are differentiated into variety of cell types:
(a) In aerial roots, the multiple epidermal cells are modified into velamen, which absorb water from the atmosphere, e. g., Orchids.
(b) Some of the cells in the leaves of grasses are comparatively very large, called bulliform or motor cells. They are hygroscopic in nature, thin-walled and contain big central vacuoles filled with water. They play an important role in the folding and unfolding of leaves. These cells develops from modification of epidermal cell and vein.
(c) Some members of Gramineae and Cyperaceae possess two types of epidermal cells : the long cells and the short cells. The short cells may be cork cells or silica cells.
♦ Cuticle and Wax: In aerial parts of the plant epidermis is covered by cuticle. The epidermal cells secrete a waxy substance called cutin, which forms a layer of variable thickness (the cuticle) within and on the outer surface of its all walls. It helps in reducing the loss of water by evaporation. Other substances deposited on the cuticle surface may be oil, resin, silicon and salts (cystoliths are crystals of calcium carbonate or calcium oxalate, e.g., Ficus. Druse and Raphides, e.g., Pistia). Thick cuticle is found in leaves of dry habita plants.
♦ Stomata: Stomata are minute apertures in the epidermis. Each aperture is bounded by two kidney shaped cells, called guard cells. In xerophytes, the stomata are sunken in grooves due to which rate of transpiration is greatly reduced (e.g. Nerium). Usually there is a large air cavity below each aperture, it is called substomatal cavity. Guard cells are surrounded by subsidiary cells or accessory cells which differ morphologically from the other epidermal cells. In monocots, e.g., Doob, Maize; guard cells are dumb bell shape.
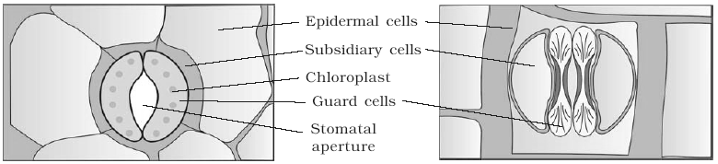
- The outer wall of guard cell is thin and the inner wall is thick .
- Chloroplast is present in guard cell, which helps in stomatal movement.
- Stomata aperture, guard cells and subsidiary cells together is refered as stomatal aperture.
♦ Trichomes : These are epidermal outgrowths present temporarily or permanently on almost all plant parts. They may be unicellular or multicellular, vary in size and shape in different species. They may be of different types: stellate hair, glandular hair, short glandular hair, urticating hair and stinging hair. The trichomes serve for checking excess loss of water and for protection.
♦ Root hairs : They are enlargements of special epiblema cells called trichoblasts and occurs in a particular zone of young root called root hair zone. They are specialised to absorb water from soil. They also hold soil particles.
Ground or Fundamental tissue system
Ground tissue system includes all the tissues of plant body except epidermal tissue system and vascular tissues. It forms the bulk of plant body. This tissue system mainly originates from ground meristem. The ground tissue constitute the following parts :
(i) Cortex: It lies between epidermis and the pericycle. The cortex is distinct in dicotyledons but not in monocotyledons, where there is no clear demarcation between cortex and pith. It is further differentiated into
(a) Hypodermis: It is collenchymatous in dicot stem and sclerenchymatous in monocot stem. It provides strength.
(b) General cortex: It consists of parenchymatous cells. Its main function is storage of food.
(c) Endodermis (Starch sheath): It is mostly single layered and is made up of parenchymatous, barrel shaped compactly arranged cells.
- The inner or transverse wall of endodermal cells have Casparian strips which has deposition of suberin.
- In roots, thick walled endodermal cells are interrupted by thin walled cells just outside the protoxylem patches. These thin walled endodermal cells are called passage cells.
- Endodermis with characteristic casparian bands is absent in woody dicot stem, monocot stem and leaves of angiosperms.
- The young stems of angiosperms show a layer with abundant starch deposition. This layer occurs in the position where endodermis would have been situated which is called as starch sheath.
- Endodermis behave as water tight dam to check the loss of water and air dam to check the entry of air in xylem elements.
- Endodermis is internal protective tissue.
(ii) Pericycle: It is a single layered or multilayered cylinder of thin-walled or thick-walled cells present between the endodermis and vascular tissues.
♦ In some cases, the pericycle is made up of many layers of sclerenchymatous cells (Cucurbita stem) or in the form of alternating bands of thin-walled and thick-walled cells (Sunflower stem).
♦ In monocot, the pericycle is made up of thin-walled parenchymatous cells which later on gives rise to lateral roots.
♦ In dicot roots the cork cambium originates in the pericycle which results in the formation of periderm.
♦ Pericycle also gives rise to a part of vascular cambium in dicot roots.
(iii) Pith or medulla: It occupies the central part in dicot stem, and monocot roots.
- It is mostly made up of parenchymatous cells.
- In dicot roots pith is completely obliterated by the metaxylem elements.
- In dicot stem, the pith cells between the vascular bundles become radially elongated and known as primary medullary rays or pith rays. They help in lateral translocation.
Vascular tissue system
The central cylinder of the shoot or root surrounded by cortex is called stele.
- The varying number of vascular bundles formed inside the stele constitute vascular tissue system.
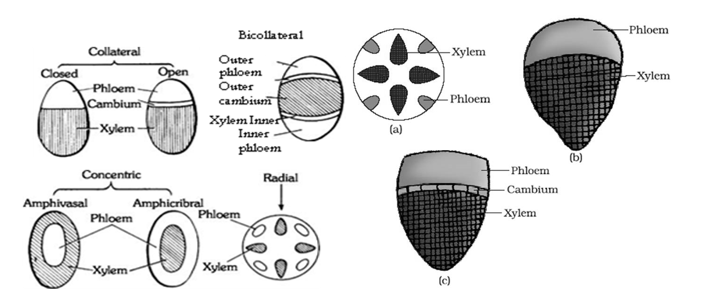
- Xylem, phloem and cambium are the major parts of the vascular bundle. Vascular bundle may be of following types :
(i) Radial: The xylem and phloem strands alternate with each other separated by parenchymatous cells. such kinds of vascular bundles are called radial and found mainly in roots.
(ii) Conjoint: A vascular bundle having both xylem and phloem together, is called conjoint. Normally the xylem and phloem occur on the same radius. They occur in stems. Such vascular bundles are of two types :
(a) Collateral: A vascular bundle in which the phloem lies towards outerside and xylem towards inner side, is called collateral, e.g., Sunflower.
▪ Collateral bundle having a cambium between xylem and phloem is said to be of the open type, e.g., Dicot stem.
▪ Collateral bundle lacking a cambium between xylem and phloem is said to be of the closed type, e.g., Monocot stem.
(b) Bicollateral: A vascular bundle having the phloem strands on both outer and inner side of xylem, is called bicollateral. e.g., Cucurbita.
(iii) Concentric: A vascular bundle in which one tissue is completely surrounded by the other, is called concentric. The concentric bundles are of two types :
(a) Amphivasal (Leptocentric): The phloem lies in the centre and remains completely surrounded by xylem. e.g., Dracaena, Yucca.
7. PRIMARY STRUCTURES IN PLANT
Difference between internal structure of root and stem
| Description | Root | Stem |
| (i) Epidermis or Epiblema | Epiblema or piliferous layer without cuticle | Epidermis usually with cuticle. |
| (ii) Hair | Unicellular | Multicellular. |
| (iii) Chlorenchyma in cortex | Absent | Usually present in young stems but absent in old stem. |
| (iv) Endodermis | Very distinct | Poorly developed or absent. |
| (v) Vascular bundle | Radial | Conjoint, collateral or bicollateral concentric. |
| (vi) Xylem | Exarch (protoxylem towards periphery) | Endarch. (protoxylem towards centre) |
Origin of Lateral roots: Lateral roots arise endogenously, i.e., from the cells inside the endodermis. They arise from pericycle cells.
8. INTERNAL STRUCTURE OF LEAF
- Like the root and stem, the leaf consists of three tissue systems, the dermal system, consisting of the upper and lower epidermis, the ground tissue system, the main photosynthetic tissue, which consists of mesophyll, and the vascular system, comprising of veins of various degrees.
- Common leaves are bifacial and are further of two types, dorsiventral and isobilateral. Unifacial leaves are cylindrical and occur in onion and garlic.
Epidermis
- The upper as well as the lower surface of the leaf is covered by a uniseriate epidermis. However, in some plants (e.g., Nerium, Ficus, etc.) the epidermis is multiseriate.
- All epidermal cells of a leaf are alike. The epidermal cells are compactly arranged and their outer walls are usually thickened. The epidermis is covered by a layer of cuticle; the thickness of the, which varies considerably and the xerophytic species have a thicker cuticle.
- In some xerophytic leaves, especially those of grasses, the epidermal cells situated in longitudinal furrows are large with thin flexible walls. These cells are said to be motor cells or bulliform cells, and they help in rolling of leaves in dry weather.
- A characteristic feature of the leaf epidermis is the presence of numerous small openings, called stomata. They occur either on both sides of the leaf (leaf is said to be amphistomatic), confined to the lower surface of the leaf (leaf is known as hypostomatic) or to the upper surface as in floating leaves of aquatic plants (leaf is called epistomatic). A stoma consists of two highly specialized epidermal cells, known as guard cells, enclosing a space. In some plants (e.g., Nerium), stomata are present in sunken cavities, called stomatal crypts.
Mesophyll
- The bulk of the internal tissue of the leaf, enclosed by the upper and lower epidermis, forms mesophyll. It is composed of thin walled parenchymatous cells containing numerous chloroplasts. The mesophyll is differentiated into palisade and spongy parenchyma in dicot leaves.
- The spongy parenchyma consists of irregular and loosely arranged cells, enclosing large intercellular spaces. These air spaces are connected with the substomatal chambers and maintain gaseous exchange with the outside through stomata.
- The palisade parenchyma is composed of more or less cylindrical and elongated cells arranged compactly with their long axis perpendicular to the epidermis.
Vascular system
- The mid rib in most dicotyledons consists of a single large collateral vascular bundle with an adaxial xylem and abaxial phloem.
The cells surrounding the vascular bundles in the leaf are mostly morphologically distinct from the mesophyll cells. These cells constitute the bundle sheath. In dicotyledons the vascular bundles are surrounded by thin walled parenchymatous cells that extend in the direction parallel to the veins. In monocotyledons, the vascular bundles are completely or partially surrounded by one or two bundle sheaths, each consisting of a single layer of cells.
Difference between dicot and monocot leaf
| Character | Dicot leaf | Monocot leaf |
| (i) Type of leaf | Dorsiventral (bifacial) | Isobilateral. |
| (ii) Stomata | Usually more on lower epidermis. | Equal on lower and upper epidermis (amphistomatic). |
| (iii) Mesophyll | Made up of two types of tissues (a) Palisade parenchyma. (b) Spongy parenchyma with large intercellular spaces | Only spongy parenchyma is present, which has very small intercellular spaces. |
| (iv)Bundle sheath | Made up of parenchyma. Just above and below the vascular bundle some parenchymatous cells or collenchymatous cells are present (upto epidermis). | Made of parenchyma, but just above and below the vascular bundles sclerenchymatous cells (upto epidermis) are found. |
| (v) Bulliform or motor cells | Absent. | Present on upper epidermis. |
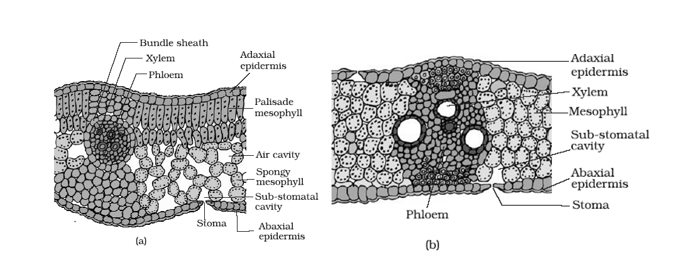
Kranz type anatomy occurs in both monocot and dicot leaves of some tropical and arid area plants.
- Kranz anatomy is characteristic feature of C4 plants.
- The mesophyll is undifferentiated and occurs in concentric layers around vascular bundles. Cells of bundle sheath possess large chloroplast.
9. INTERNAL STRUCTURE OF STEM
- The part of the axis of the plant, which is usually ascending and aerial and also bears leaves and reproductive structures is called The stem has conspicuous nodes and internodes and differs fundamentally from the root in its vascular structure.
- Vascular bundles are conjoint, collateral or bicollateral or even concentric, open or closed.
- A transverse section of young stem shows the following structures:
Epidermis
- It is represented by the outermost layer of the stem, which is protective in function. It is made up of a single layer of closely packed rectangular cells with relatively thin primary walls. The epidermis is covered on the outer surface by a layer of cuticle. The continuity of epidermis is broken by the presence of stomata. Multicellular hairs (trichomes) are frequently found on the epidermis.
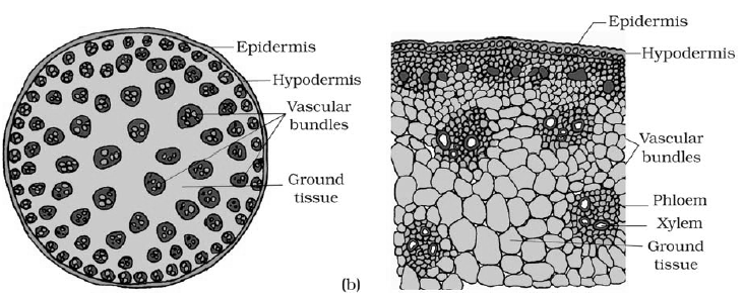
Fundamental (ground) tissue system
- In dicotyledonous stems the ground tissue is clearly differentiated into cortex and pith. But in monocotyledonous stems there is no such distinction in the ground tissue.
- In dicotyledonous stems the cortex is usually differentiated into three distinct regions, an outer collenchymatous hypodermis, a middle parenchymatous general cortex and an inner endodermis.
- Hypodermis: It consists of a few layers of collenchymatous cells immediately below the epidermis. It usually forms a continuous ring in cylindrical stems, but in angular stems, it occurs in patches (e.g., Cucurbita).
- General Cortex: It is parenchymatous with polygonal, round or oval cells having intercellular spaces. Chloroplasts may be present in the outer layers of the cortex. Latex tubes, resin ducts, crystals of calcium oxalate and reservoirs of waste products are frequently present in the cortex.
- Endodermis: It is the innermost limiting layer of the cortex, which separates vascular tissue from the cortex. It is made up of vertically elongated cells, which appear barrel shaped in cross section. These cells, however, do not develop casparian thickening like those of the root endodermis. The cells of this layer contain abundant starch and hence it is also called starch sheath.
- Pericycie: The layer of cells, which occur inner to the endodermis and outside the vascular tissue form pericycle. It is usually made up of closely fitting fibres, which occur in isolated patches or form a complete cylinder. It is multilayered and may be parenchymatous (e.g., most of the dicots) or sclerenchymatous (e.g., cucurbits) or both (e.g., Asteraceae).
- Pith: It is the ground tissue present in the centre of the stem. It is made up of large parenchymatous cells with distinct intercellular spaces. Laticifers, secretory cells, sclereids, idioblasts, crystals of various kinds and other ergastic substances are present in the pith. The outer part of the pith is sometimes distinct in having smaller cells with thicker walls. Such a morphologically distinct outer pith is called perimedullary zone or medullary sheath.
- Medullary rays: These are narrow, vertically elongated bands of parenchymatous cells which extend transversely from the pith towards the periphery. The first rays, formed as a result of the growth of the apical meristem, are known as primary medullary rays.
Vascular tissue system
- The vascular tissue is in the form of distinct vascular bundles. In dicotyledonous stems, vascular bundles are arranged in a ring. They are conjoint, collateral, bicollateral or concentric and open. Such stele in which vascular bundles are arranged in ring is called In monocot stems vascular bundles are scattered and such stele is called atactostele. Usually the outer bundles are smaller and gradually become larger as they pass inward. The bundles are collateral and closed.
- Xylem: It consists of vessels, tracheids, xylem parenchyma and xylem fibres. The protoxylem lies towards the inner side of the bundle (endarch condition). The cells constituting the protoxylem are smaller in diameter than those of metaxylem in the form of ‘Y’. The two metaxylem vessels are situated at the tips of the arms of ‘Y’ and the other two at the base of ‘Y’. The protoxylem often disintegrates to form a schizolysigenous cavity (protoxylem lacuna).
- Phloem: It consists of sieve tubes, companion cells, phloem parenchyma and phloem fibres. In monocots, the phloem, which is situated in between the arms of ‘Y’, does not have phloem parenchyma.
- Cambium: It is a band of thin walled cells, which lie between the phloem and xylem: The cells constituting cambium are thin walled and rectangular in cross section. They have prominent nuclei and dense cytoplasm. During secondary growth, by rapid division in cambial cells, secondary phloem and secondary xylem are formed. Cambium is absent in monocots.
Difference Between dicot and monocot stem
|
Characters |
Monocotyledonous Stem |
Dicotyledonous stem |
|
(i) Epidermis
|
Present, cells comparatively smaller and without hair. |
Present, cells larger and with hair. |
|
(ii)Hypodermis |
Sclerenchymatous (non-green) |
Collenchymatous (green) |
|
(iii)Cortex
|
Absent, but ground tissue is present from hypodermis to the centre of stem. |
Made up of several layers of parenchymatous tissue. |
|
(iv)Endodermis
|
Absent
|
One layered, starchy sheath which is usually not well differentiated. |
|
Characters |
Monocotyledonous Stem |
Dicotyledonous stem |
|
(v)Pericycle
|
Absent
|
Made up of one or more layers of parenchymatous and sclerenchymatous cells. |
|
(vi)Medullary rays
|
Absent
|
Found in between vascular bundles |
|
(vii)Pith (Medulla)
|
Absent
|
Abundant, made up of parenchymatous cells |
|
(viii)Vascular bundles
|
▪ Scattered ▪ Conjoint, Collateral and closed ▪ Larger towards centre ▪ Oval ▪ Bundle sheath present ▪ Phloem parenchyma absent ▪ Xylem vessels either Y or V shaped |
▪ Vascular bundles in a ring ▪ Conjoint, collateral and open ▪ All of same size ▪ Usually wedge—shaped ▪ Bundle sheath absent ▪ Phloem parenchyma present ▪ Xylem vessels more radial |
ANATOMICAL FEATURES OF PLANT PARTS
10. INTERNAL STRUCTURE OF ROOT
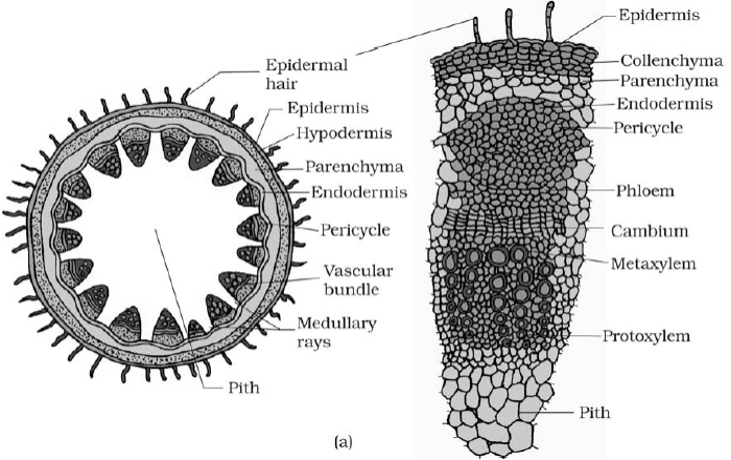
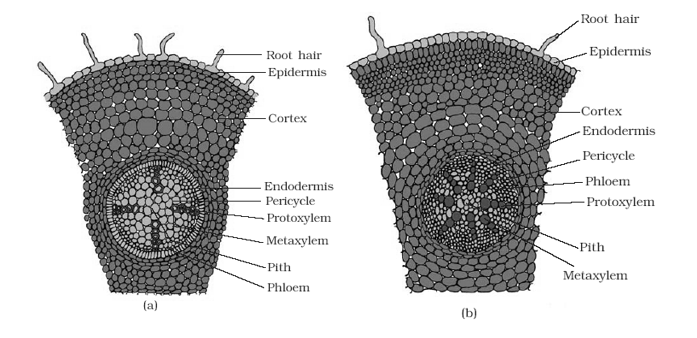
- Root is part of the plant axis, which normally grows below the soil surface and functions as an anchoring and absorbing organ. It has a comparatively simple internal organization than that of a stem due to the absence of nodes and internodes and lateral appendages.
- The arrangement of tissues from the periphery to the centre of a primary root is as follows:
Epiblema
- This is the outermost layer of tightly placed thin walled and non-cutinized cells. The outer wall of the cells of epiblema gives rise to unicellular root hair. These hairs help in absorption of water from the soil.
- Epiblema is usually uniseriate, but in aerial roots of some orchids and epiphytic aroids multi seriate epidermis, called velamen, is present.
Cortex
- The root has a histologically simple cortex usually composed of thin walled isodiametric parenchymatous cells with many intercellular spaces. The cortical cells are usually devoid of chloroplasts but they store starch.
- Some roots (e.g., Iris) develop a specialized layer beneath the epiblema, called exodermis. It arises from subepidermal layers of the cortex. The cells of exodermis have suberized cell walls.
Endodermis
- Endodermis delimits cortex from the stele. It is composed of closely packed living cell: which are characterized by the presence of casparian strips in their radial walls.
- Some cells of endodermis, usually the ones which lie opposite to protoxylem, remain thin walled.
- These cells, called passage cells or transfusion tissue allow radial diffusion of water.
Pericycle
- Pericycle occurs beneath the endodermis and it forms the outer boundary of the primary vascular cylinder of the root. It is usually a single layer of cells.
Vascular System
- The vascular bundles of roots are radial with alternating strands of xylem and phloem separated by parenchymatous conjunctive tissue. The xylem is exarch.
- In dicot roots, the number of xylem bundles usually vary from 2 to 6 and in monocot roots, it usually ranges from 6 to 20. Depending on the number of xylem bundles, roots are designated as monarch (with one xylem bundle), diarch (two bundles), triarch (three bundles), tetrarch (four bundles), pentarch (five bundles) or polyarch (many bundles).
Pith
- Pith occupies a very small area in the centre of the root. It consists of thin walled parenchymatous cells with intercellular spaces.
- In many dicot roots, due to the development of metaxylem vessels in the centre, pith is obliterated.
Difference between dicot and monocot root
| Character | Dicot Root | Monocot Root |
| (i) Pericycle | Gives rise to secondary roots and lateral meristem | Gives rise to lateral roots only |
| (ii) Vascular bundles | Diarch to hexarch | Hexarch to polyarch |
| (iii) Cambium | Develops at the time of secondary growth | Absent |
| (iv) Pith | Absent or poorly developed | Abundant and fully developed |
| (v) Secondary growth | Takes place | Does not take place |
| Narrow cortex. Endodermis is less thickened and Cortex wide. Casparian ships are visible casparian ships are more prominent. | Cortex wide. Casparian strip are visible only in young root. Later on endodermal cells become highly thickened. | |
| Conjuctive tissue present | Absent |
11. SECONDARY GROWTH
The increase in thickness or girth due to the activity of the cambium and the cork cambium is known as secondary growth.
Secondary growth in stem
On the basis of the activities of cambium and cork-cambium, secondary growth in stem can be discussed under the following heads :
(i) Activity of cambium (ii) Activity of cork-cambium
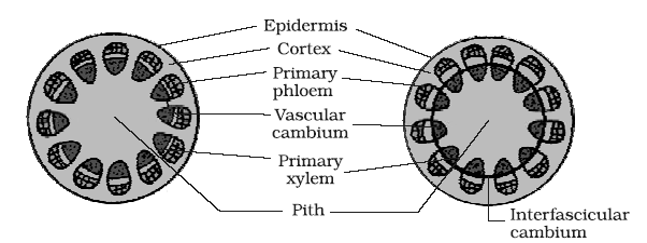
(i) Activity of cambium: The vascular cambium in between xylem and phloem is called intrafascicular or fascicular cambium which is primary in origin. At the time of secondary growth the parenchymatous cells of medullary rays between the vascular bundles become meristematic and form strip of cambium called as interfascicular cambium which is secondary in origin. Both inter and intrafascicular cambium joins together and form cambium ring which is partly primary and partly secondary in origin. By anticlinal divisions the circumference of the cambium increase. By periclinal division cambium produces the secondary xylem and phloem tissues on innerside and outerside, the amount of sec. xylem produced is 8-10 times greater than secondary. phloem. The cambium has two types of cells :
(a) The fusiform initials which are elongated form fibres, sieve cells, sieve tubes, and tracheids.
(b) Ray initials which produce parenchyma cells of the rays in wood and phloem.
♦ Ray initials are much shorter than fusiform initials.
♦ Certain cells of cambium form some narrow bands of living parenchyma cells passing through secondary xylem and secondary phloem and are called secondary medullary rays.
♦ These provide radial conduction of food from the phloem; and water and mineral salts from the xylem.
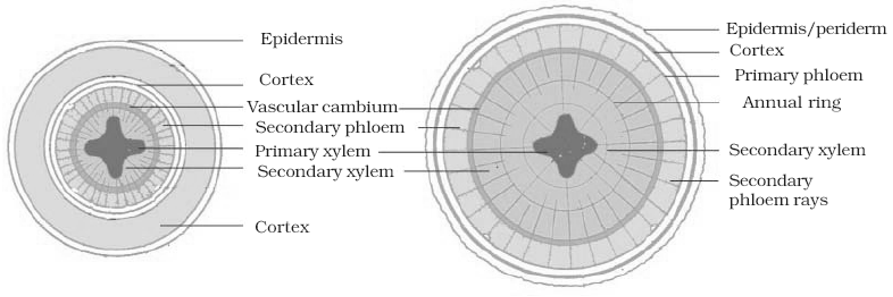
- Annual rings: Activity of cambium is not uniform in those plants which grow in the regions where favourable climatic conditions (spring or rainy season) alternate regularly with unfavourable climatic conditions (cold water or dry hot summer).
- In temperate climates, cambium becomes more active in spring and forms greater number of vessels with wider cavities; while in winter it becomes less active and forms narrower and smaller vessels.
- The wood formed in the spring is known as spring wood and they are formed in the dry summer or cold winter as autumn wood or late wood. Both autumn and spring wood constitute a growth or annual ring.
- In one year only one growth ring is formed. Thus by counting the number of annual rings in the main stem at the base we can determine the age of a tree. This branch of science is known as dendrochronology.
- Growth rings are distinct or sharply demarcated in the plants of temperate regions, where as in tropical climate (near equator) they are not distinct or sharply demarcated.
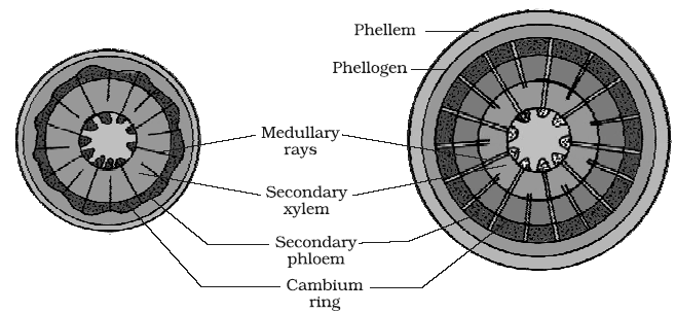
(ii) Activity of cork cambium: Cork cambium or phellogen develops from outer layer of cortex. It produces secondary cortex or phelloderm on innerside and cork or phellem on outerside.
- The cells of phellem are dead, suberized and impervious to water.
- Cells of phelloderm are thin walled, living and store food.
- Phellem, phellogen and phelloderm is collectively called as periderm. Periderm is secondary protective tissue.
- Due to pressure of secondary xylem, epidermis ruptures and cortex is largely lost after two or three years of secondary growth.
(a) Bark: All tissues lying outside the active vascular-cambium are collectively known as bark.
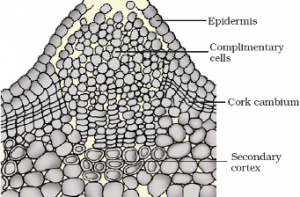
- This includes secondary phloem and periderm.
- When cork-cambium appears in the form of a complete ring, it is known as ring bark, e.g., Betula (Bhojpatra).
- If the cork cambium occurs as separate strips and the resulting bark appears in the form of scales, such a bark is known as scaly bark. e.g., Eucalyptus, Psidium guava.
- The outermost layer of bark is dead and called as rhytidome.
(b) Lenticels : These are aerating pores formed in the cork through which gaseous exchange takes place.
- They are formed as a result of the action of phellogen.
- A lenticel appears as a scar or protrusion on the surface of the stem and consists of a radial row of thin-walled cells, known as complementary cells or filling tissue. They are found in old dicot stem.
(c) Cork: It consists of dead cells with thick walls, heavily impregnated with suberin.
- These cells are compactly arranged in radial rows without intercellular spaces.
- Cork is impervious to water and prevents its loss from the plant surface.
- It also protects the inner tissues from the attack of fungi and insects.
(d) Heart wood and sap wood: In old trees, secondary wood is differentiated into a centrally situated darker, harder wood called the heart wood or duramen.
- Cells of heart wood are physiologically inactive (almost dead). Outer light-coloured zone is called the sap wood or alburnum which is physiologically active.
- Dark colour of heart wood is due to the deposition of tannins, resins, gums, essential oils, etc. in the cell walls and cell cavities.
- The water conduction takes place through sap wood.
- During the conversion of sap wood into heartwood, the most important change is development of tyloses in the heart wood. Tyloses are ballon like structures, which develop from xylem parenchyma. These tyloses block the passage of xylem vessels, therefore, also called as tracheal plug.
- The heart wood is commercially used as wood. When the plant is made hollow, it will not die because the water conduction takes place through sap wood. The heart wood is well developed in Morus alba (Mulberry). The heart wood is absent in Populus and Salix plant. As a tree grows older, thickness of heartwood increases and sap wood remains same.
Secondary growth in dicot roots
Vascular bundles in dicot roots are radial, exarch and mostly triarch.
- Vascular cambium is formed secondarily from conjuctive parenchyma cells lying just below each phloem strand.
- Thus the number of cambium strips formed equals the number of phloem strands.
- The cells of pericycle lying outside the protoxylem also become meristematic to form part of strips of cambium. These cambial strips join the first formed cambium strips to form complete but wavy ring of vascular cambium. This cambium ring produced secondary xylem on inner side and secondary phloem on the outer side.
- In roots, the growth rings are not distinct because there is no seasonal variation in the soil.
- From the outer layers of pericycle arises the phellogen, which cuts phellem (cork) on the outer side and secondary cortex or phelloderm toward the inner side.